Guest blogger: Anna Lundquist (ESCI and PHIL double-major, class of 2027)
Diatoms are microscopic, single-cellular algae that build their skeletons (called frustules) out of silica, aka glass! They are extremely abundant, living basically anywhere they can find water and sunlight, in both freshwater and marine environments; in fact, they produce over half of the Earth’s oxygen. They are also very sensitive to environmental change, and preserve well, making them excellent for all kinds of paleoecological analysis.
This summer I attended the 63rd annual 2026 Iowa Lakeside Lab Ecology of Systematics of Diatoms course, lovingly nicknamed Diatom Camp, a month-long summer class that gave me the opportunity to spend a month at the University of Iowa’s field station in Okoboji, Iowa.

MacBride Lab, where we spent our class (and microscope!) time
There, I spent time learning about diatom taxonomy and how they can be used for paleolimnological interpretation, as well as spending lots of time in the field. The campus is located right next to a lake on 147 acres of prairie. Through the course, I got to explore prairie, fen, lake, river, and bog environments, under the microscope and in the field! The most important material I will be working with for my Senior Independent Study are lake core samples, so getting the opportunity to take and work with fresh lake cores and surface samples in a field setting in Iowa was an especially exciting experience.

Collecting a sediment sample from West Lake Okoboji using the Ponar surface sampler

Exploring Silver Lake Fen, a calcareous wetland filled with unique muck and marl pools
Over the course of the class, we got to work with samples not just from Iowa, but also marine estuaries in Florida, and streams from Minesota’s North Shore, which is currently facing an infestation of invasive didymo, or rock snot, the only diatom with a common name. Rock snot forms slimy brown mats of mucilage, a mucus-like substance that diatoms secrete to help them anchor to a substrate, on the bottom of otherwise clean and low nutrient streams. This is a nuisance to both human and non-human visitors to the streams, with didymo choking out everything else living in the streams. In class, we looked at samples from North Shore streams, to help assess the spread of didymo, as well as looking at the other kinds of diatoms present.
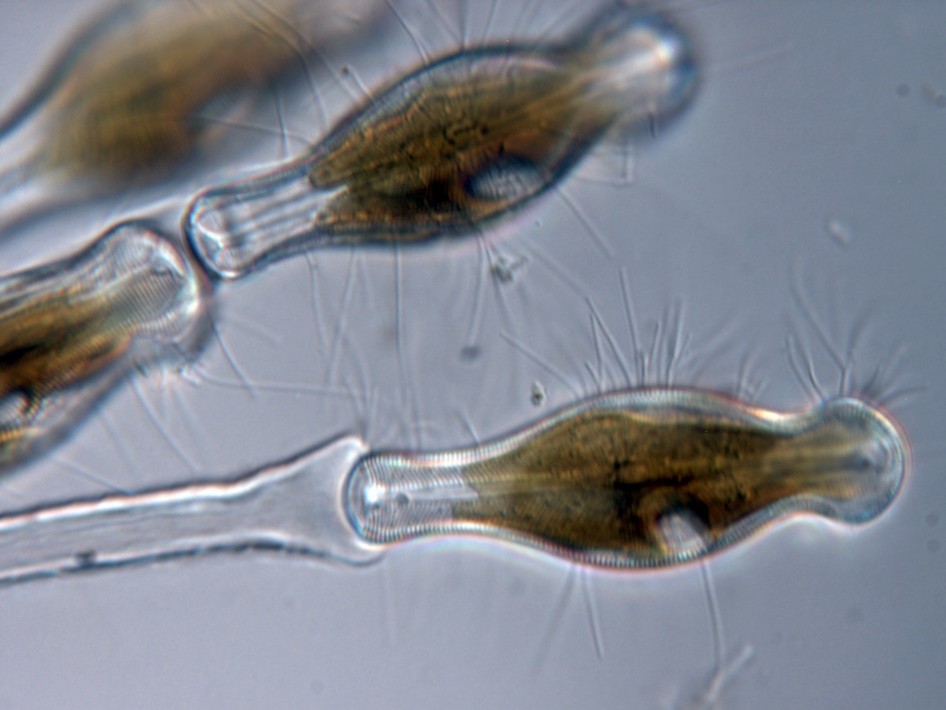
A living sample of didymo, showing the mucilage stalk. Photo courtesy of Sarah Spaulding via diatoms.org.
As the only undergraduate student in the class, I also was able to meet graduate students working on a diverse range of projects in geology, paleontology, and environmental science. Over the course of the class, I got to learn about their research as well as getting to work alongside them as peers and friends.

Our class photo in front of MacBride Lab
I was taking this course to support my Independent Study, which will investigate the effects of climate change and human activity on two small lakes in the eastern Sierra Nevada using diatom stratigraphy. Over the next year, I hope to use my newly gained diatom knowledge to track and compare palaeoecological changes in these two lakes using core samples. I also hope to examine surface sediment samples to map out a biogeographical profile of the sediment characteristics and diatom communities living in different areas of both lakes.

Looking in the microscope
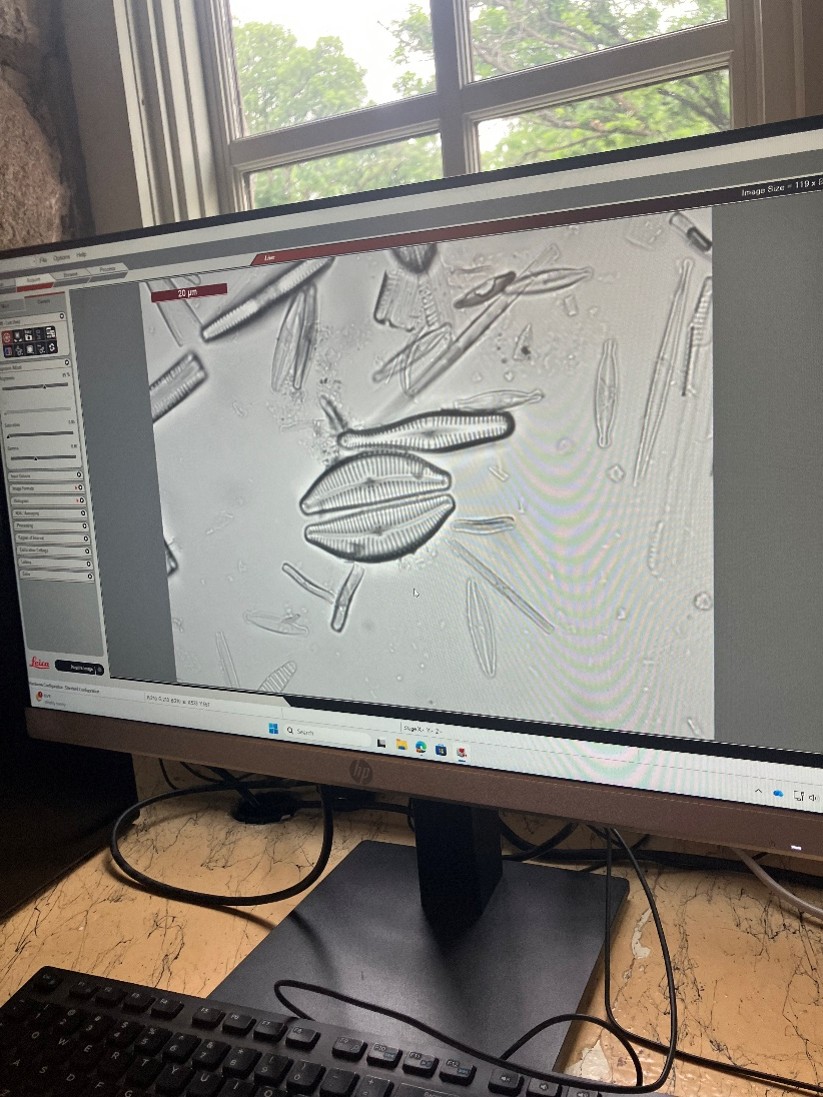
The view through the scope!
My experience in this course would not have been possible without the support of Wooster’s Copeland fund and Earth Science department’s Davis fund, and I am so grateful to have had this opportunity!






 Yellow cedar (left) and redcedar (right)
Yellow cedar (left) and redcedar (right)
 Perseverance Lake.
Perseverance Lake.




















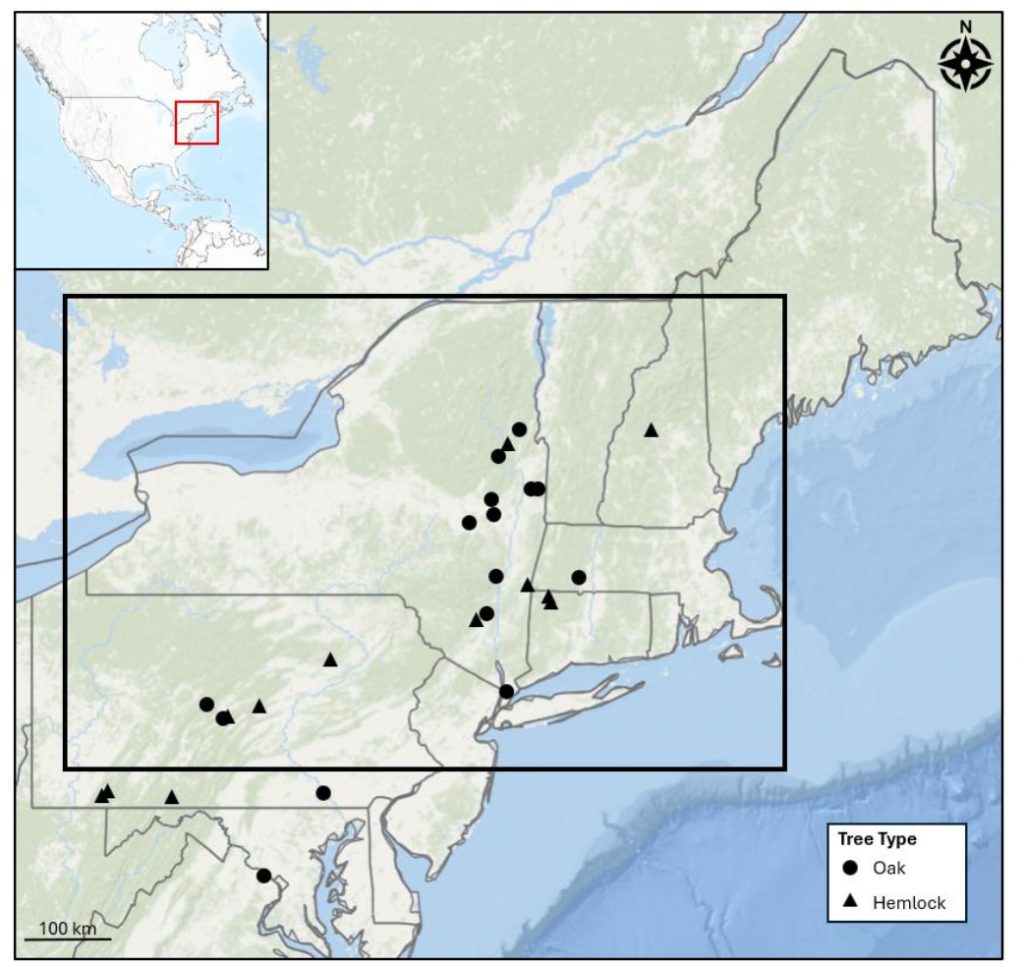
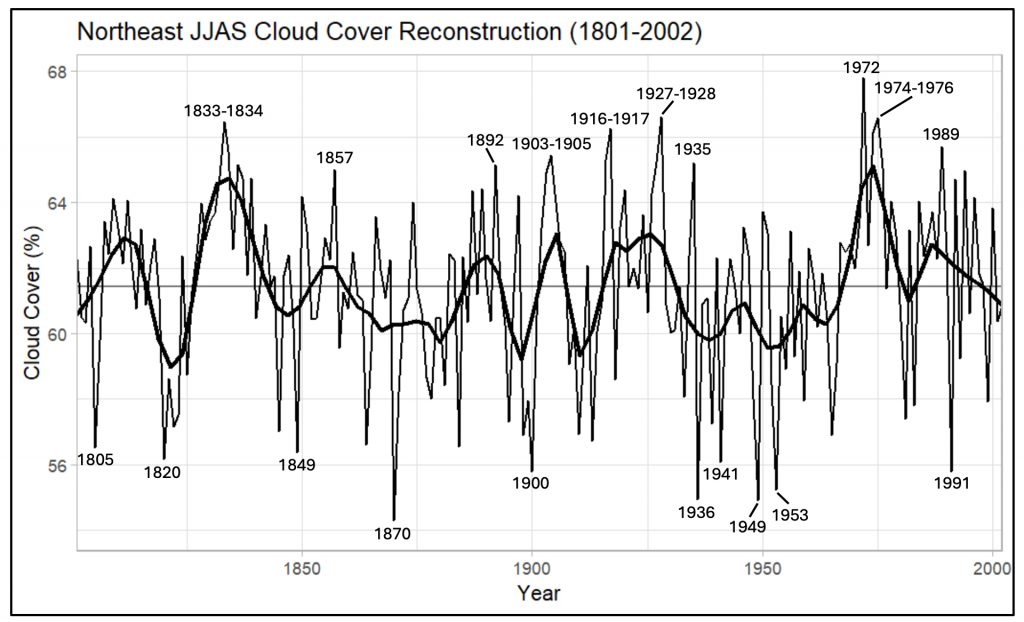
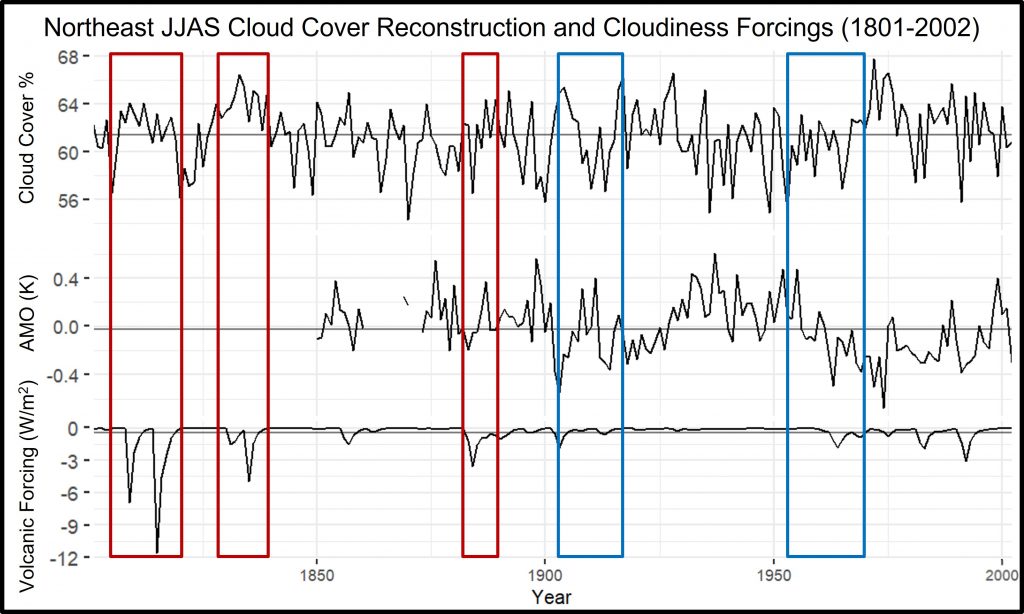



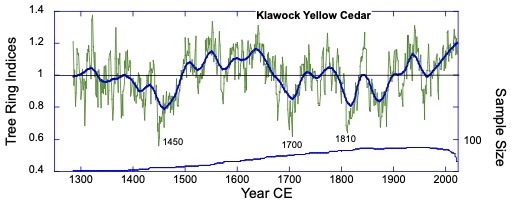
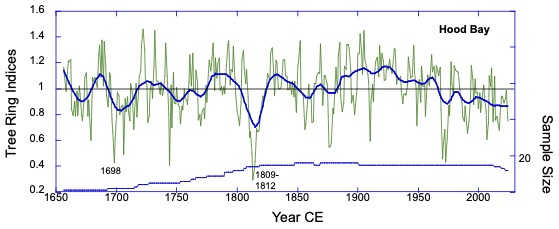
 Background: The collaboration started remotely in the summer of 2021 – the summer of 2022 was a visit to Kake, then Hoonah (2023), Klawock (2024) and then in 2025 Angoon. Throughout the years, all the AYS (Alaska Youth Stewards) groups continued coring trees and sending samples to The Wooster Tree Ring lab and meeting online.
Background: The collaboration started remotely in the summer of 2021 – the summer of 2022 was a visit to Kake, then Hoonah (2023), Klawock (2024) and then in 2025 Angoon. Throughout the years, all the AYS (Alaska Youth Stewards) groups continued coring trees and sending samples to The Wooster Tree Ring lab and meeting online.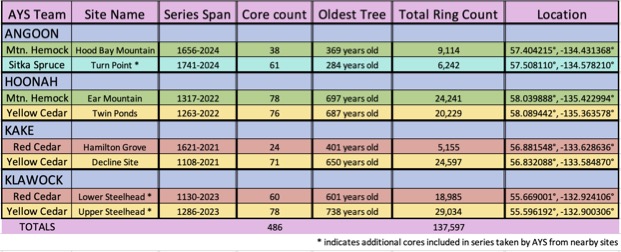
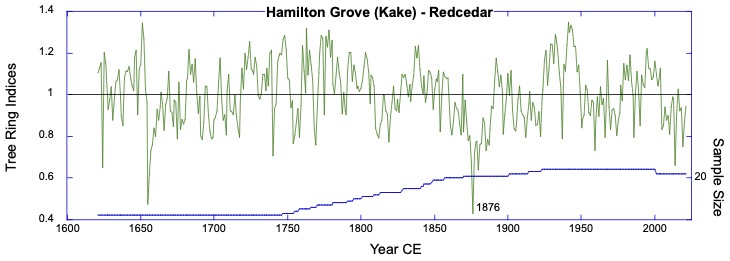


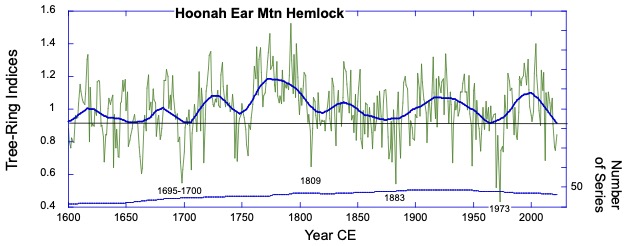
 Alaska yellow cedar near Hoonah stripped between 2001 and 2006 CE (Common Era). The year of stripping was determined by coring within and outside of the scar and then taking the difference between the total rings in each core. The stripping is sustainable and not detected as a decline in growth in the ring-width series.
Alaska yellow cedar near Hoonah stripped between 2001 and 2006 CE (Common Era). The year of stripping was determined by coring within and outside of the scar and then taking the difference between the total rings in each core. The stripping is sustainable and not detected as a decline in growth in the ring-width series.