


ICELAND – Team Iceland is investigating the formation of subglacial pillow lavas on the Reykjanes Peninsula in southwest Iceland.
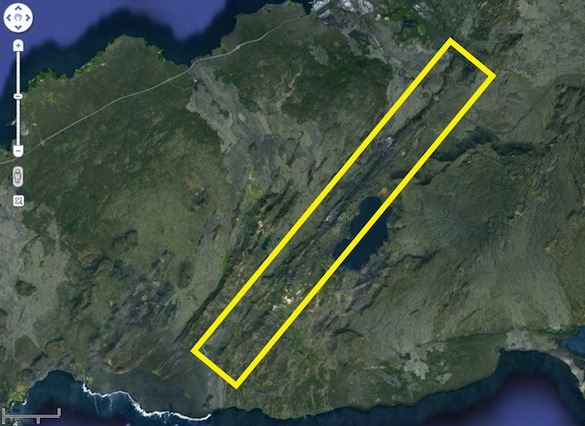
Google Earth image showing the elongated pillow ridge that we are working on. The pillow ridge erupted along a fissure system when the peninsula was glaciated.
Quarries along the ridge expose the internal structure of the subglacial deposits, revealing complex sequences of pillow lavas, intrusions, and fragmental units.
One of the active quarries graciously allowed us to use their break room for a morning overview.

Ben Edwards discusses the geological goals of the project. Photo Credit: Jim Ciarrocca
We spent most of the rest of the day working in the inactive part of the quarry.

Team Iceland examines the base of a wall of pillow lavas and discusses the formation of hyaloclastite, the glassy fragmental material that spalls off the pillow rims.

As we explored the walls, we found a lava cave. Alex Hiatt (’14) snapped a photo of the hibernating lava bears for Dr. Wilson.

Aleks (’14, Dickinson) uses a GPS and a laser range finder to “shoot” the quarry walls.

Adam Silverstein (’16) points out features on the wall for Aleks to shoot.
The day ended with some reconnaissance work in the active parts of the quarry after the workers had left.

Team Iceland poses with a a fantastic columnar jointed basalt they found on their reconnaissance investigation.


























