


 I’ve spent much of my career investigating marine sclerobionts through time. A sclerobiont is an organism that lives on or within a hard substrate. Among marine sclerobionts are oysters that encrust cephalopod shells, barnacles attached to boat hulls, and clams that bore into coral reef. My favorite historical examples are the sclerobiont communities inhabiting the abundant brachiopod shells in the Cincinnati Group (Upper Ordovician) of Ohio, Indiana and Kentucky. These diverse assemblages, like those on the brachiopod shell above, are fun to study because they record fossilized organisms in their original positions (in situ) on these hard substrates. This means we can plot out relative timing of community development by mapping overlapping encrusters and borings that cut through the resulting encruster stratigraphy.
I’ve spent much of my career investigating marine sclerobionts through time. A sclerobiont is an organism that lives on or within a hard substrate. Among marine sclerobionts are oysters that encrust cephalopod shells, barnacles attached to boat hulls, and clams that bore into coral reef. My favorite historical examples are the sclerobiont communities inhabiting the abundant brachiopod shells in the Cincinnati Group (Upper Ordovician) of Ohio, Indiana and Kentucky. These diverse assemblages, like those on the brachiopod shell above, are fun to study because they record fossilized organisms in their original positions (in situ) on these hard substrates. This means we can plot out relative timing of community development by mapping overlapping encrusters and borings that cut through the resulting encruster stratigraphy.
But what if occasionally encrusters were removed from these assemblages without leaving any traces of their existence? This is the dilemma described in a new Historical Biology paper by myself and colleagues Caroline Buttler and Olev Vinn. Below is the abstract with some of the critical figures —
Abstract — The abundant shells and hardgrounds in the Cincinnatian Group (Upper Ordovician, Katian) of the upper midwestern United States were commonly encrusted and bored by a variety of organisms. Numerous studies of these sclerobiont communities have provided valuable data for models of ecological succession, symbiosis, space and food resource competition, and taphonomy. An underlying assumption of this work is that most of the skeletal encrusters have remained in place, firmly attached to their hard substrates. This is especially the case with the most common encrusters, thick trepostome bryozoan and cystoporate skeletons, on their most common substrates, flat strophomenide brachiopods. We present evidence here, though, that these bryozoans were often dislodged from their brachiopod hosts, leaving no evidence of their attachment other than horizontal borings in semi-relief from organisms that excavated tunnels (Trypanites and Palaeosabella) at the interface of the brachiopod shell and attaching bryozoan. Similar borings are found on the bases of dislodged bryozoans and in bryoimmured mollusc external moulds. These borings along the bryozoan attachment surfaces caution us that there are significant numbers of missing skeletal encrusters on these hard substrates.
 Figure 1. External ventral valves of Cincinnatian strophomenide brachiopods showing Trypanites and Palaeosabella borings in semi-relief (‘unroofed’), location C/W-153 (A, B, E, F), and attachment surfaces (undersides) of trepostome bryozoans showing Trypanites and Palaeosabella borings in semi-relief (‘unfloored’), location C/W-148 (C, D). (A) Unroofed borings in a variety of directions in the brachiopod valve; specimen #CW153–1. (B) Unroofed borings perpendicular to the brachiopod commissure; specimen #CW153–2. (C) Mostly Palaeosabella borings distinguished by distal clavate terminations; specimen #CW148–1. (D) Mostly Trypanites borings with cylindrical forms; specimen #CW148–2. (E) External ventral valve; specimen #CW153–3. (F) Internal ventral valve; specimen #CW153–4.
Figure 1. External ventral valves of Cincinnatian strophomenide brachiopods showing Trypanites and Palaeosabella borings in semi-relief (‘unroofed’), location C/W-153 (A, B, E, F), and attachment surfaces (undersides) of trepostome bryozoans showing Trypanites and Palaeosabella borings in semi-relief (‘unfloored’), location C/W-148 (C, D). (A) Unroofed borings in a variety of directions in the brachiopod valve; specimen #CW153–1. (B) Unroofed borings perpendicular to the brachiopod commissure; specimen #CW153–2. (C) Mostly Palaeosabella borings distinguished by distal clavate terminations; specimen #CW148–1. (D) Mostly Trypanites borings with cylindrical forms; specimen #CW148–2. (E) External ventral valve; specimen #CW153–3. (F) Internal ventral valve; specimen #CW153–4.
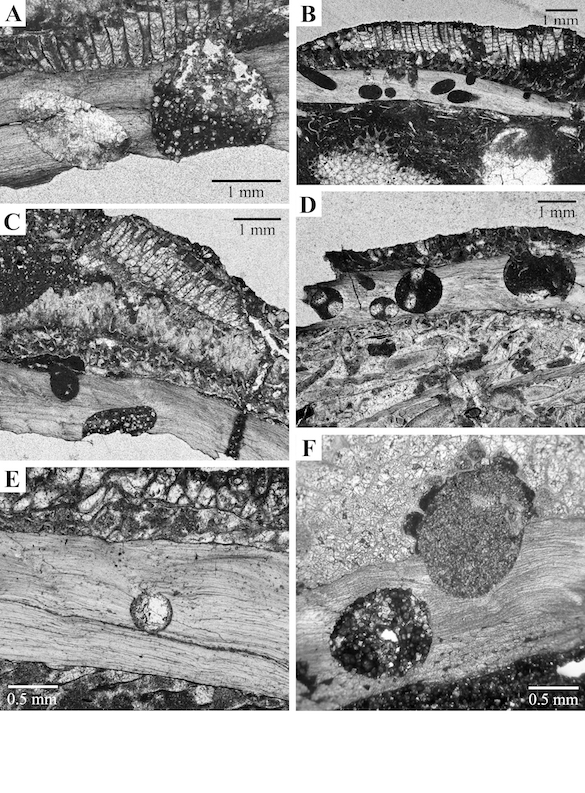 Figure 2. Thin sections (A–E) Location C/W- 153 and acetate peel (F) Location C/W-148, perpendicular to brachiopod valves encrusted by bryozoans colonies, all extensively bored. (A) Borings infilled with dolomite rhombs (right) and calcite (left); specimen #CW153–5. (B) Borings infilled with micrite, some within brachiopod shell, and other cutting through shell and encrusting bryozoans; specimen #CW153–6. (C) Borings infilled with dolomite rhombs and micrite; specimen #CW153–5. (D) Borings showing two ghosts inside, suggesting that the soft-bodied organism was U-shaped along the axis of the boring; specimen #CW153–7. (E) Small boring within brachiopod shell infilled with calcite; specimen #CW153–7. (F) Brachiopod valve encrusted with a trepostome bryozoan with two cylindrical borings parallel to the shell: the one on the right cuts through the brachiopod and encrusting bryozoan; specimen #CW148–3.
Figure 2. Thin sections (A–E) Location C/W- 153 and acetate peel (F) Location C/W-148, perpendicular to brachiopod valves encrusted by bryozoans colonies, all extensively bored. (A) Borings infilled with dolomite rhombs (right) and calcite (left); specimen #CW153–5. (B) Borings infilled with micrite, some within brachiopod shell, and other cutting through shell and encrusting bryozoans; specimen #CW153–6. (C) Borings infilled with dolomite rhombs and micrite; specimen #CW153–5. (D) Borings showing two ghosts inside, suggesting that the soft-bodied organism was U-shaped along the axis of the boring; specimen #CW153–7. (E) Small boring within brachiopod shell infilled with calcite; specimen #CW153–7. (F) Brachiopod valve encrusted with a trepostome bryozoan with two cylindrical borings parallel to the shell: the one on the right cuts through the brachiopod and encrusting bryozoan; specimen #CW148–3.
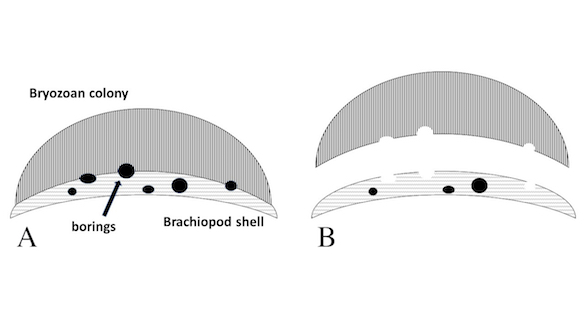 Figure 3. Diagram showing the proposed process for forming unroofed borings in brachiopods and unfloored borings in encrusting bryozoans. (A) Cross-section of brachiopod valve with encrusting bryozoan attached; the borings are perpendicular to the cross-section plane; note borings that excavate both the brachiopod shell and the overlying encrusting bryozoan. (B) Same cross-section with the encrusting bryozoan detached, leaving unfloored borings in the bryozoan base and unroofed borings in the brachiopod valve.
Figure 3. Diagram showing the proposed process for forming unroofed borings in brachiopods and unfloored borings in encrusting bryozoans. (A) Cross-section of brachiopod valve with encrusting bryozoan attached; the borings are perpendicular to the cross-section plane; note borings that excavate both the brachiopod shell and the overlying encrusting bryozoan. (B) Same cross-section with the encrusting bryozoan detached, leaving unfloored borings in the bryozoan base and unroofed borings in the brachiopod valve.
The implications of this taphonomic process that can remove encrusting bryozoans from shelly substrates include the important reminder that critical information is always missing from our paleontological data sets. In this case even the simple observation that a particular fossil brachiopod is not encrusted does not mean it wasn’t previously encrusted before burial.
Reference cited:
Wilson, M.A., Buttler, C.J. and Vinn, O. 2024. Traces of missing encrusters: borings reveal sclerobiont taphonomy in the Upper Ordovician (Katian) of the Cincinnati region, USA. Historical Biology (in press). https://doi.org/10.1080/08912963.2024.2312402